Your shopping cart is empty!

This may be of several types.
The mother cells divide and the daughter cells are produced, which become new plants. This is exclusive type of reproduction in Pleurococcus, some desmids, diatoms, Euglena, etc.
The plant body breaks into several parts or fragments and each such fragment develops into an individual. This type of vegetative reproduction is commonly met within filamentous forms, e.g., Ulothrix, Spirogyra, etc. The fragmentation of colonies also takes place in several blue green algae, e.g., Aphanocapsa, Aphanothece, Nostoc, etc.
When the trichomes break in small pieces of two or more cells, such pieces are called ‘hormogones’. Each hormogone develops into a new plant, e.g., Oscillatoria, Nostoc, etc.
They are thick-walled hormogones, and produced in somewhat drier conditions.
Certain special structures of thalli are formed which help in vegetative reproduction. The well known propagula of Bryopsis, Sphacelaria and Nereocystis are good examples.
Such thread-like vegetative bodies develop in the case of Chara, which help in reproduction.
Usually these bodies are rounded and filled up with abundance of starch. Each body may give rise to a new plant, e.g., Chara.
Such special star-shaped, starch filled bodies give rise to new plants frequently reported from Chara.
Small bud-like structures. Usually develop on the rhizoids of Chara are called bulbils. Each such bulbil may develop into a new plant.
In most of the Chlorophyceae members, the Akinetes are developed. Usually the protoplast of each cell converts in a single akinete. Sometimes they are formed in chains. Each akinete may develop into a new plant, e.g., Oedogonium, Ulothrix, etc.
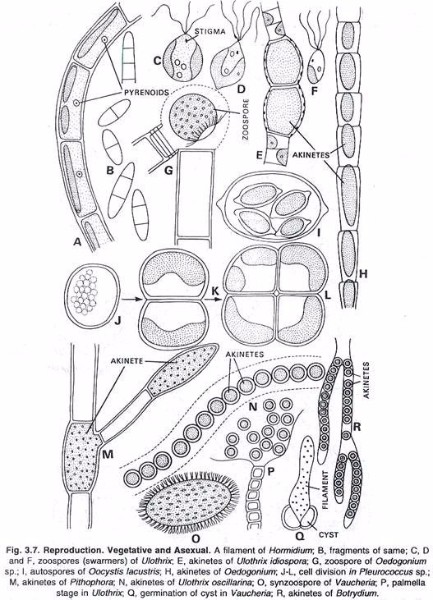
Usually the protoplast of a cell divides into several protoplasts and thereafter they escape from the mother and develop into new plants. (See Fig. 3.7).
The zoospores are formed from certain older cells of the filaments. The cytoplasm divides to form zoospores which are escaped from the mother cell. They are always formed in favourable conditions. The zoospores are always motile. They may be (i) biflagellate, (ii) tetraflagellate, (iii) stephanokontean type of zoospores, e.g., Oedogoniales and (iv) compound zoospores, e.g., Vaucheriaceae.
When motile phase of zoospores is eliminated, the bodies are called aplanospores. The aplanospores develop in unfavourable conditions. Each such spore is surrounded by a wall.
Actually they are very thick-walled aplanospores and develop only in adverse conditions, e.g., Pediastrum, Vaucheria.
Here the successive generations of divided cells are gelatinized and a thick mucilaginous envelope develops, e.g., Chlamydomonas, Ulothrix, etc.
They are just like aplanospores except that they are smaller in size. They resemble in shape to mother cell except in size. Each autospore gives rise to a new plant. Such autospores are reported from many Chlorococcales.
In many blue green algae and Bacillariophyceae, the endospores are formed within the cells. On the approach of favourable conditions, each endospore develops in a new individual.
In many members of Bacillariophyceae, such auxospores are produced. Each develops in a new plant.
They are found in the carposporophytes of red algae (Rhodophyceae). Each such spore develops in a new individual.
These spores are not formed within the sporangia. They are found in Rhodophyceae.
These spores develop within monosporangia. Each spore gives rise to a new plant, e.g., many members of Rhodophyceae (Bangia, Porphyra, Porphyridium, etc).
Such spores are reported from many members of Rhodophyceae. Each spore develops into a new plant.
They are found in Xanthophyceae and Bacillariophyceae where they act as perennating bodies.
In many Volvocales and Chlorococcales, the daughter colonies are developed asexually, e.g., Volvox, Hydrodictyon, Pediastrum, etc.
In the aseptate filaments of Vaucheria, the protoplast divides into several parts, several hypnospores or cysts are produced and the whole filament looks like an algal form ‘Gongrosira’.
They are produced in many Bacillariophyceae.
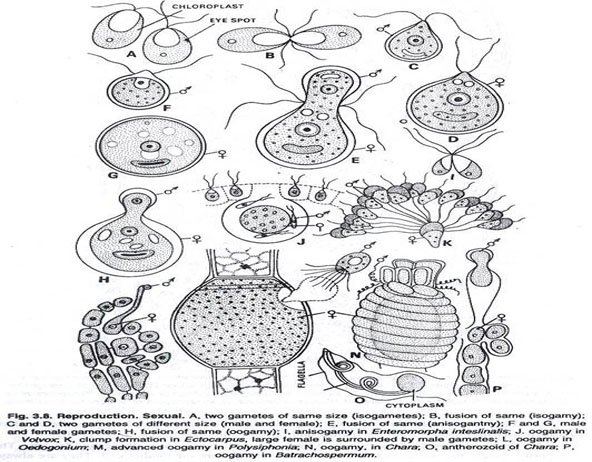
It is greatly advanced method of reproduction and not known in Myxophyceae (blue green algae). There are two main types, i.e., (i) isogamy and (ii) heterogamy.
The fusion of similar motile gametes is found in many species. Usually the gametes taking part in fusion come from two different individuals or filaments, sometimes these gametes come from two different cells of the same filament. Thousands of gametes come and aggregate in clumps. (See Fig. 3.8).
The fusion of dissimilar gametes is called heterogamy. There are variations of it.
The motile gametes taking part in fusion may either differ in size (morphological anisogamy) or physiological behaviour (physiological anisogamy).
In this case, the male antherozoid fuses with the female egg. This fusion may be of primitive type as found in Cylindrocapsa, or advanced type as in Oedogonium, Vaucheria, Chara, Polysiphonia, etc.
It implies the fusion of two non-flagellate amoeboid gametes (aplanogametes). They are morphologically similar but physiologically dissimilar, e.g., order Conjugales.
In fresh water algae, the sexual reproduction is best means of perennation because it is followed by the formation of thick-walled zygote or oospore.
(a) The sexual reproduction takes place after considerable accumulation of food material and the climax of vegetative activity is over.
(b) The bright light is the major factor for the production of the gametes.
(c) A suitable pH value is required.
(d) The optimum temperature is necessary.
The female gametes convert into zygotes without fusion. The resultants are called azygospores or parthenospores and the phenomenon ‘parthenogenesis’, e.g., Spirogyra, Oedogonium and many others.
In this phenomenon, the fusion of the daughter protoplasts or of the divided nuclei of a cell without liberation takes place. This process is known in many diatoms and colourless dinoflagellates.
We have algae available on https://seatechbioproducts.com/marine-algae-micro-and-macro-seaweeds
